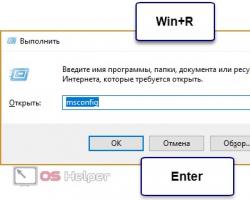
Лекция 11
Основным механизмом противовирусного иммунитета являются иммунные клеточные реакции, осуществляемые Т-эффекторами, а основную роль в этих клеточных реакциях играют Т-киллеры, которые распознают зараженную клетку в организме и вызывают ее цитолиз. В результате организм освобождается от клеток, продуцирующих инфекционное вирусное потомство.
Индуцированные вирусным антигеном Т-лимфоциты приобретают свойства распознавать вирусный антиген, находящийся на поверхности зараженных клеток. Вирусные детерминанты, распознаваемые Т-лимфоцитами, сходны или идентичны детерминантам, выявляемым В-лимфоцитами на поверхности зараженных клеток. Однако в отличие от В-лимфоцитов, Т-лимфоциты распознают только те зараженные клетки, на поверхности которых вирусный антиген сцеплен с клеточными антигенами главного комплекса гистосовместимости - HLA у человека.
Т-лимфоциты обладают высокой специфичностью и могут, например, различать клетки, которые заражены вирусами гриппа типа А или гриппа типа В. Однако специфичность Т-лимфоцитов в отношении вирусов относительна и варьирует у разных таксономических групп.
Существенным фактором в противовирусном иммунитете являются макрофаги. Они принимают участие в иммунной стимуляции, распознавании антигена, регуляции пролиферации и дифференцировки лимфоцитов. Кроме того, они являются активными помощниками в разрушении и удалении из организма неродственных антигенов. Цитотоксическая активность макрофагов имеет неспецифический характер и проявляется на ранних стадиях инфекционного процесса.
В противовирусном иммунитете имеют значение и другие факторы клеточного иммунитета, такие как активность естественных киллеров и зависимая от антител цитотоксичность, обусловленная неиммунными лимфоидными клетками.
Клеточный иммунитет, как указывалось, играет более важную роль при вирусных инфекциях, нежели гуморальный иммунитет. Лишь часть вирусов (в частности, пикорнавирусы) быстро разрушают пораженные ими клетки, большинство же вирусов не вызывают немедленной их гибели, а онкогенные вирусы, наоборот, вызывают пролиферацию пораженках клеток. Поэтому зараженные клетки становятся мишенью для цитолитического действия Т-эффекторов, естественных киллеров и макрофагов. Цито-литическое действие имеет место при всех вирусных инфекциях и поэтому должно быть отнесено вместе с интерфероном (см. далее) к основным факторам, способствующим выздоровлению организма от вирусной инфекции.
Особенности иммунитета при некоторых вирусных инфекциях. Продолжительность и прочность иммунитета варьирует при разных вирусных инфекциях. Так, при оспе, кори, паротите иммунитет является весьма прочным. При других вирусных инфекциях иммунитет не столь стоек и продолжителен и поэтому возможны повторные заболевания. Это, по-видимому, относится к некоторым парамиксовирусам (респираторно-синцитиальный вирус), а также к риновирусам, хотя повторное заражение может объясняться заражением другими серологическими типами риновирусов.
Своеобразны особенности иммунитета при гриппе. Перенесенная инфекция создает стойкий иммунитет: так, первая волна гриппа в 1977-1978 гг., вызванная вирусом H1N1, циркулировавшим в 1957 г., характеризовалась тем, что заболели почти исключительно лица моложе 20 лет, родившиеся после 1957 г. Таким образом, повторные заболевания гриппом одного и того же серотипа связаны не с нестойкостью иммунитета, а с антигенным дрейфом двух поверхностных вирусных белков. О прочности иммунитета при гриппе свидетельствует феномен антигенной доминанты или «первородного антигенного греха». При повторных заболеваниях гриппом наряду с появлением антител к вирусу, вызвавшему заболевание, стимулируется рост антител к вирусу, с которым произошла первая встреча данного индивидуума. Этот феномен был широко использован для «серологической археологии» - выяснения, какие вирусы гриппа циркулировали в прошлом, до их открытия.
Многие вирусы персистируют в организме, несмотря на наличие антител. Например, аденовирусы могут длительно персистировать в миндалинах, вирусы герпеса могут длительно и даже пожизненно сохраняться в нервных клетках чувствительного ганглия тройничного нерва или в дорзальных ганглиях. При ряде персистентных вирусных инфекций причиной персистенции является недоступность вируса для циркулирующих в крови антител. Некоторые вирусы способны распространяться из клетки в клетку без выхода во внеклеточное пространство. Например, вирусы герпеса могут проникать из одной клетки в другую по цитоплазматическим мостикам. Многие вирусы (вирусы парагриппа человека, кори, респираторно-синцитиальный и др.) вызывают слияние соседних клеток и распространяются путем формирования симпласта или синцития. Возможно проникновение вируса и субвирусных компонентов в дочерние клетки при клеточном делении. Существует несколько способов «ускользания» вирусов от иммунологического надзора: 1) подавление фагоцитоза; 2) угнетение Т- и В-системы; 3) особая локализация вируса в организме, защищающая его от действия им-муноцитов. В результате создаются условия для распространения вируса в организме и его персистенции. Нарушение функции лимфоцитов имеется при большинстве вирусных инфекций. Вирусы гриппа, кори, полиомиелита, герпеса, ротавирусы и особенно вирус СПИД угнетают иммунные реакции Т-лимфоцитов, препятствуют их стимуляции. Вирус СПИД вызывает, деструкцию Т-хелперов. Вирусы герпеса - возбудители ветряной оспы и опоясывающего герпеса, цитомегалии, инфекционного мононуклеоза - приводят к увеличению абсолютного и относительного количества Т-супрессоров. Активацию су-прессоров вызывает вирус клещевого энцефалита.
Таким образом, приобретенный иммунитет после перенесенной вирусной инфекции может быть разным: в одних случаях он защищает от повторных заболеваний на многие годы или на всю жизнь; в других случаях утрачивается через несколько лет и даже месяцев, в связи с чем возможны повторные заболевания; в третьих случаях иммунитет не предотвращает персистирование вируса в организме и появление периодических рецидивов.
ИММУНОПАТОЛОГИЧЕСКИЕ РЕАКЦИИ
В иммунологии иммунопатологическими реакциями называют такие иммунологические феномены, которые приводят к повреждению органов и тканей хозяина и одновременно направлены против возбудителя. Однако при вирусных инфекциях наряду с иммунопатологическими реакциями, наблюдающимися и при других инфекциях (образование иммунных комплексов, аутоантител и др.), встречаются и необычные феномены, которые также можно отнести к иммунопатологии.
Сохранение инфекционного вируса в иммунном комплексе и макрофагах. При взаимодействии вирусов с антителами могут формироваться иммунные комплексы, в которых вирусы сохраняют инфекционную активность. Обычно это имеет место при использовании недостаточной концентрации антител, однако избыток антител не всегда оказывает дополнительный инактивирующий эффект при ряде инфекций. Длительная циркуляция в организме таких иммунных комплексов приводит, во-первых, к постоянному инфицированию чувствительных клеток, во-вторых, к антигенной стимуляции вирусспецифических иммунокомпетентных клеток. В результате формируются новые иммунные комплексы, содержащие инфекционный вирус. Образующиеся иммунные комплексы фиксируются на клетках, содержащих рецепторы к Fc-фрагменту иммуноглобулина, и в результате создаются условия для прикрепления и проникновения в клетки вируса.
Увеличение количества связавшегося с клеткой вируса, находящегося в составе иммунных комплексов, объясняется следующими причинами.
1. Клеточные рецепторы для ряда вирусов не могут обеспечить такого эффективного проникновения в клетку.
2. Агрегированный вирус труднее проникает в клетку, чем мономерная вирусная частица, окруженная антителами.
3. Антитела защищают вирус от протеолитической деградации клеточными ферментами.
Феномен парадоксального усиления репродукции вирусов при использовании недостаточных концентраций антивирусных антител присущ многим вирусам: альфа- и флавивирусам, буньявирусам, рабдовирусам, реовирусам. Особенно четко этот феномен проявляется при заражении вирусами макрофагов: связанные с антителами вирусные частицы лучше размножаются в макрофагах по сравнению со свободными. Наиболее четко феномен проявляется при использовании вируса денге (флавивирус).
Макрофаги способны фагоцитировать многие вирусы. Однако не все фагоцитированные вирусы разрушаются ферментными системами макрофагов; незавершенный фагоцитоз в ряде случаев может не предотвращать развитие инфекции, а становиться ее источником. Такая инфекция может протекать как в острой, так и хронической форме. Примером устанавливающейся персистенции при взаимодействии вируса и макрофагов является экспериментальная цитомегалия.
Иммунная деструкция зараженных клеток. Клеточные мембраны могут разрушаться гуморальными факторами - лимфотоксинами, которые синтезируются лимфоцитами. Лимфотоксины оказывают неспецифическое ферментативное действие вблизи секретирующих их клеток. Однако основным механизмом деструкции зараженных клеток является цитотоксическое действие Т-лимфоцитов. Вирус-специфические Т-киллеры появляются вскоре после заражения, через 1-3 суток. Способность Т-лимфоцитов разрушать зараженные клетки может приводить не только к защитному эффекту и выздоровлению от инфекции, но и к иммунопатологическим реакциям в результате поражения органов и тканей. Защитное или повреждающее действие Т-лимфоцитов зависит от стадии инфекции, в течение которой они действуют. Если разрушение зараженных клеток происходит на ранних стадиях инфекции, гибель немногочисленных зараженных клеток, находящихся, в основном, у входных ворот инфекции, не приведет к нарушению гомеостаза, и наступит выздоровление. Напротив, при действии Т-лимфоцитов на более поздних стадиях инфекции, когда в результате распространения вируса в организме повреждены клетки многих органов и тканей, иммунный цитолиз может привести к некомпенсируемым нарушениям жизненно важных функций организма и усугубить инфекционный процесс.
В ряде случаев специфические антитела могут воздействовать на клетки в отсутствии комплемента (при некоторых иммунодефицитных состояниях, поражении нервных клеток, недоступных для комплемента и т. д.). Антитела в отсутствие литического действия комплемента приводят к уменьшению выхода вирусных белков на клеточную поверхность, в результате чего может развиться внутриклеточная персистенция вирусных компонентов.
Единый механизм, лежащий в основе защитного и повреждающего действия иммунных Т-лимфоцитов, предполагает обязательное участие иммунопатологического компонента в патогенезе любой вирусной инфекции, и иммунопатологию можно рассматривать как обязательную плату за выздоровление при вирусных инфекциях. При разных инфекциях вирусной этиологии соотношение защитного и повреждающего действия Т-лимфоцитов существенно варьирует.
Аутоиммунные антитела. Деструкция зараженных вирусом клеток в процессе инфекции приводит к появлению антигенно измененных клеточных структур, которые воспринимаются организмом как чужеродные и вызывают формирование гуморальных и клеточных факторов иммунитета, способных взаимодействовать с антигенами нормальных клеток. Конформационная перестройка молекулы антигена, взаимодействующего с антителом, также является причиной образования аутоантител против собственных иммуноглобулинов. В результате возникают аутоиммунные реакции. В их патогенезе важную роль играет нарушение сосудистой проницаемости под действием иммунных комплексов. В результате происходит антигенная стимуляция элементов лимфоидной ткани, синтез аутоантител и формирование ауто-Т-лимфоцитов, разрушающих ставшие чужеродными клеточные антигены. Аутоиммунные процессы часто приводят к развитию осложнений при вирусных инфекциях. С аутоиммунными процессами связано, например, возникновение орхита как осложнения при вирусном паротите, обусловленного повышением проницаемости кровеносных и лимфатических сосудов тестикулярной ткани; возникновение миокардита при инфекции, вызванной вирусами Коксаки. У больных хроническим гепатитом В обнаруживаются клетки с цитотоксической активностью к гепатоцитам, которая реализуется в присутствии антител против специфического липопротеина печени, находящегося на поверхности гепатоцитов.
Иммунокомплексная патология. Образование иммунных комплексов при взаимодействии вируса с антителами является важным механизмом, обеспечивающим выздоровление и формирование противовирусного иммунитета. Однако иммунные комплексы могут оказывать не только защитное, но и повреждающее действие на организм. Иммунокомплексная патология широко распространена при вирусных инфекциях и играет значительно большую роль в их патогенезе по сравнению с другими инфекционными и неинфекционными болезнями.
В образовании иммунных комплексов участвуют преимущественно антитела класса IgG, однако их формирование может происходить и с участием IgM- и IgA-антител. Так, IgM- и IgA-антитела в составе иммунных комплексов были обнаружены в клубочковых отложениях почек при алеутской болезни норок, инфекционном мононуклеозе, гепатите В. Формирование иммунных комплексов происходит как в жидкой среде, так и на поверхности зараженных вирусом клеток. Может происходить связывание антител с вирусными гликопротеидами, расположенными на поверхности зараженных клеток, с последующим освобождением иммунных комплексов во внеклеточное пространство. Такие иммунные комплексы образуются при кори. Размер, растворимость и биологическая активность комплексов зависят от отношения антигена к антителу и их относительной концентрации. Иммунные комплексы склонны к агрегации и вторичному связыванию различных молекул: компонентов комплемента, антител к ним, антиглобулинов, антиидиотипических антител.
Судьба иммунных комплексов и их биологическая активность в организме могут быть различными. Они могут взаимодействовать с клетками иммунной системы путем связывания с рецепторами к Fc-фрагменту иммуноглобулинов, при этом аффинитет к рецепторам иммунных комплексов повышен по сравнению с иммуноглобулином. Взаимодействие с рецептором ведет к активации клеток, секреции биологически активных веществ, которые увеличивают проницаемость сосудов, активизируют свертывающую систему крови и т. д. Иммунные комплексы могут изменить гуморальный и клеточный ответ путем взаимодействия с В- и Т-клетками, усиливать или подавлять активность лимфоцитов.
Иммунные комплексы могут циркулировать в кровотоке и в межтканевой жидкости. Крупные иммунные комплексы быстро выводятся из циркуляции с участием системы мононуклеарных фагоцитов. Мелкие иммунные комплексы могут фиксироваться в стенках сосудов или мембранах почечных клубочков. Иммунные комплексы, находящиеся в межтканевых пространствах, плохо выводятся и вызывают локальное воспаление ткани. Укрупнение иммунных комплексов, их сорбция на стенках сосудов и тканях вызывают повреждение органов и тканей и обусловливают «болезни иммунных комплексов», патогенез которых связан с развитием гломерулонефрита. и впервые разработан при сывороточной болезни.
Элементы иммунокомплексной патологии встречаются при большинстве вирусных инфекций и играют значительную роль в их патогенезе. Наиболее изучена иммунокомплексная патология при гепатите В, герпетической инфекции, геморрагической лихорадке денге, подостром склерозирующем панэнцефалите. В продромальном периоде гепатита В возникают васкулиты и артриты, обусловленные циркулирующими иммунными комплексами, содержащими HBs-антиген и анти-НВв-антитела, и гломерулонефриты, связанные с гранулярными отложениями иммунных комплексов в ткани почек.
В ядрах гепатоцитов при хроническом гепатите В регулярно обнаруживают как вирусные антигены, так и антитела класса IgG, которые проникают в клетку в результате нарушения проницаемости клеточных мембран. Основным антигенным компонентом является HBs-антиген, однако в иммунных комплексах обнаруживаются НВс- и НВе-антигены. При геморрагической лихорадке денге иммунные комплексы обусловливают усиленную репродукцию вируса путем взаимодействия с Fc-рецепто-рами моноцитов и активацию системы комплемента. При инфекционном мононуклеозе, лимфоме Беркитта и назофарингеальной карциноме обнаружены иммунные комплексы, содержащие аутоантитела. У больных цитомегалией детей обнаружены циркулирующие иммунные комплексы и иммунные комплексы в мембранах почечных клубочков. При ряде вирусных инфекций иммунокомплексная патология лежит в основе патогенеза. Так, гибель мышей, внутриутробно зараженных вирусом лимфоцитарного хориоменингита, наступает впоследствии от иммунокомплексного гломерулонефрита; та же причина гибели наблюдается при алеутской болезни норок. При других вирусных инфекциях иммунокомплексная патология явно не выражена, однако она также играет роль в патогенезезаболевания. Например, при полиовирусной инфекции формирование иммунных комплексов, фиксирующихся в стенках кровеносных сосудов, приводит к нарушению их проницаемости и способствует проникновению вирусных частиц через гематоэнцефалический барьер.
Сохранение инфекционной активности вируса в составе иммунных комплексов является одной из основных причин возникновения хронических форм вирусных инфекций. При этом создается порочный круг: длительно текущий патологический процесс наносит ущерб репарационным системам гомеостаза организма, что, в свою очередь, приводит к созданию условий для персистенции вируса или его компонентов.
ИНТЕРФЕРОН
А. Айзекс и Ж. Линдеман в 1957 г. обнаружили, что клетки, зараженные вирусом, вырабатывают особое вещество, угнетающее размножение как гомологичных, так и гетерологичных вирусов, которое они назвали интерфероном. В дальнейшем было показано, что существует много интерферонов и поэтому следует говорить о системе интерферона. Если иммунная система обеспечивает белковый гомеостаз и через него устраняет чужеродную генетическую информацию, то система интерферона непосредственно воздействует на чужеродную генетическую информацию, устраняя ее из организма на клеточном уровне, и тем самым обеспечивает нуклеиновый гомеостаз. Система интерферона тесно взаимодействует с иммунной системой.
Интерфероны являются белками с молекулярной массой, колеблющейся у разных интерферонов от 22х10 3 (мышиный интерферон) до 94х10 3 (интерферон форели).
Интерфероны закодированы в генетическом аппарате клетки. Гены для человеческого фибробластного интерферона располагаются во 2-й, 9-й и длинном плече 5-й хромосомы, а ген, регулирующий транскрипцию - в коротком плече той же хромосомы. Ген, детерминирующий восприимчивость к действию интерферона, локализован в 21-й хромосоме. Ген для ά-интерферона располагается в 9-й хромосоме, для γ-интерферона - в 11-й хромосоме.
Система интерферона не имеет центрального органа, так как способностью вырабатывать интерферон обладают все клетки организма позвоночных животных, хотя наиболее активно вырабатывают его клетки белой крови.
Интерферон спонтанно не продуцируется интактными клетками и для образования его нужны индукторы, каковыми могут быть вирусы, бактериальные токсины, экстракты из бактерий и грибов, фитогемагглютинины, синтетические вещества - поликарбоксилаты, полисульфаты, декстраны, но наиболее эффективными индукторами интерферона являются двунитчатые РНК: двунитчатые вирусные РНК и двунитчатые синтетические сополимеры рибонуклеотидов (поли-ГЦ, поли-ИЦ) и др. Индукция интерферона происходит вследствие дерепрессии его генов.
Типы интерферонов. Известны три типа интерферонов человека: ά-интерферон, или лейкоцитарный интерферон, который продуцируется лейкоцитами, обработанными вирусами и другими агентами; β-интерферон, или фибробластный интерферон, который продуцируется фибробластами, обработанными вирусами и другими агентами. Оба эти интерферона принадлежат к типу 1. Более сильный γ-интерферон, или иммунный интерферон, принадлежит к типу 2. Имеется несколько подтипов ά-интерферона, и общее число их у человека доходит до 25. Сравнительная характеристика интерферонов человека приведена в табл. 14. Активность интерферонов измеряется в международных единицах (ME). Одна единица соответствует количеству интерферона, которое ингибирует репродукцию вируса на 50%.
При индукции интерферонов синтезируется два и более его типов. Так, при индукции интерферона на лимфобластах образуется 87% лейкоцитарного и 13% фибробластного интерферона, при индукции интерферона на фибробластах имеют место обратные соотношения. Между тремя типами интерферонов могут существовать синергические взаимодействия.
Свойства интерферонов. Интерфероны обладают видотканевой специфичностью. Это означает, что интерферон человека действует только в организме человека, но неактивен в организме других биологических видов. Конечно, барьеры видовой специфичности не абсолютны: интерферон человека проявляет некоторую активность в тканях человекообразных обезьян, а куриный интерферон в организме близких видов семейства куриных. Однако активность интерферона в гетерогенных организмах резко
снижается. Поэтому можно заключить, что интерфероны, появившиеся у позвоночных, эволюционировали вместе с хозяевами. Интерферон является относительно устойчивым белком и хорошо переносит кислую среду (рН 2,2), что используется для выделения его и очистки. Антигенные свойства интерферонов мало выражены, в связи с чем антитела к нему удается получить лишь после многократных иммунизации.
Интерфероны не обладают специфичностью в отношении вирусов и действуют угнетающе на репродукцию различных вирусов, хотя разные вирусы обладают неодинаковой чувствительностью к интерферону. Чувствительность к нему обычно совпадает с индуцирующей активностью к интерферону. Наиболее часто применяемыми индукторами интерферона и тест-вирусами для его титрования являются рабдовирусы (вирус везикулярного стоматита), парамиксовирусы, тогавирусы. Продукция интерферона зависит также от характера применяемых клеток. Существуют клетки, дефектные по нескольким генам интерферона.
Интерфероны оказывают антивирусное, противоопухолевое, иммуномодулирующее и многие другие действия. Наиболее изучено их антивирусное действие, и именно на вирусных моделях выяснены биологические и другие свойства интерферонов.
Интерферон оказывает противоопухолевое действие при парентеральном введении в больших дозах, связанное с подавлением им цитопролиферативной активности. Добавление интерферона к культуре нормальных клеток сопровождается уже через 2 ч угнетением в них синтеза ДНК. При вирусиндуцированных опухолях интерферон угнетает репродукцию онковирусов и одновременно подавляет цитопролиферативную активность.
Интерферон является регулятором различных механизмов иммунного ответа, оказывая стимулирующее или угнетающее действие на иммунные реакции.
Механизм действия интерферона. Интерферон связывается с клеточными рецепторами, находящимися на плазматической мембране, что служит сигналом для дерепрессии соответствующих генов. В результате индуцируется синтез особой протеинкиназы, которая присутствует в следовых количествах во всех клетках млекопитающих и активируется низкими концентрациями двунитчатой РНК, а в зараженных вирусами клетках - вирусными репликативными комплексами.
Протеинкиназа фосфорилирует ά-субъединицу инициирующего фактора трансляции eIF-2, и фосфорилирование блокирует активность инициирующего фактора. В результате иРНК, связанная с инициирующим комплексом, не может связаться с большой рибосомальной субъединицей, и поэтому ее трансляция блокируется. Инициирующий фактор eIF-2 в одинаковой степени необходим для трансляции как клеточных, так и вирусных иРНК, однако преимущественно блокируется трансляция вирусных иРНК, связанных с вирусными двунитчатыми РНК-структурами, в результате локальной активации протеинкиназы.
В обработанных интерфероном клетках индуцируется синтез фермента - синтетазы, которая катализирует 2,5-олигоадениловую кислоту, переключающую действие клеточных нуклеаз на разрушение вирусных иРНК. Таким образом, вирусные иРНК подвергаются разрушению нуклеазами. Блокирование интерфероном стадии инициации трансляции и разрушение иРНК обусловливают его универсальный механизм действия при
инфекциях, вызванных вирусами с разным генетическим материалом.
Применение интерферонов. Интерфероны применяются для профилактики и лечения ряда вирусных инфекций. Их эффект определяется дозой препарата, однако высокие дозы интерферона оказывают токсическое действие. Интерфероны широко применяются при гриппе и других острых респираторных заболеваниях. Препарат эффективен на ранних стадиях заболевания, применяется местно, например путем закапывания или введения с помощью ингалятора в верхние дыхательные пути в концентрациях до 3-10 4 - 5-10 4 ед. 2-3 раза в день. При конъюнктивитах интерферон применяют в виде глазных капель. Интерфероны оказывают терапевтическое действие при гепатите В, герпесе, а также при злокачественных новообразованиях. При этих заболеваниях назначают более высокие концентрации. Препарат применяется парентерально - внутривенно и внутримышечно в дозе 10 5 ед. на 1 кг массы тела. Более высокие дозы оказывают побочное действие (повышение температуры, головная боль, выпадение волос, ослабление зрения и т. д.). Интерферон может вызвать также лимфопению, задержку созревания макрофагов, у детей - тяжелые шоковые состояния, у больных с сердечно-сосудистыми заболеваниями - инфаркт миокарда. Очистка интерферона значительно снижает его токсичность и позволяет применять высокие концентрации. Очистка осуществляется с помощью аффинной хроматографии с использованием моноклональных антител к интерферону.
Генноинженерный интерферон. Генноинженерный лейкоцитарный интерферон получают в прокариотических системах (кишечной палочке). Биотехнология получения интерферона включает следующие этапы: 1) обработка лейкоцитарной массы индукторами интерферона; 2) выделение из обработанных клеток смеси иРНК; 3) получение суммарных комплементарных ДНК (кДНК) с помощью обратной транскриптазы; 4) встраивание кДНК в плазмиду кишечной палочки и ее клонирование; 5) отбор клонов, содержащих гены интерферона; 6) включение в плазмиду сильного промотора для успешной транскрипции гена; 7) экспрессия гена интерферона, т. е. синтез соответствующего белка; 8) разрушение прокарио-тических клеток и очистка интерферона с помощью аффинной хроматографии. Получены высокоочищенные и концентрированые препараты интерферона, которые испытываются в клинике.
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Особенности противовирусного иммунитета
Иммунную защиту макроорганизма при вирусных инфекциях осуществляет противовирусный иммунитет. Его особенности обусловлены двумя формами существования вируса: внеклеточной и внутриклеточной. Основными факторами, обеспечивающими противовирусный иммунитет, являются специфические антитела, Т-киллеры, интерферон и сывороточные ингибиторы вирусных частиц. Специфические противовирусные антитела способны взаимодействовать только с внеклеточным вирусом, внутриклеточные структуры прижизненно для них недоступны. Антитела нейтрализуют вирусную частицу, препятствуя ее адсорбции на клетке-мишени, инфицированию и генерализации процесса, и обеспечивают иммунный фагоцитоз «маркированных» вирусных частиц. Специфические антитела также связывают вирусные белки и нуклеиновые кислоты, которые попадают в межклеточную среду и секреты после разрушения зараженных вирусами клеток.
Клетки, инфицированные вирусом и приступившие к его репликации, экспрессируют (представляют) вирусные белки на поверхности цитоплазматической мембраны в составе молекул антигенов гистосовместимости -- HLA I класса. Изменение структуры этих антигенов гистосовместимости служит сигналом для активации Т-киллеров. Последние специфически распознают клетки макроорганизма, зараженные вирусом и приступившие к биосинтезу его компонентов, и уничтожают их. Мощным противовирусным действием обладает интерферон. Он не влияет непосредственно на внеклеточный и внутриклеточный вирус, а адсорбируется на мембране клеток и индуцирует ферментные системы, подавляющие синтез компонентов вируса. Сывороточные ингибиторы неспецифически связываются с вирусной частицей и нейтрализуют ее, тем самым препятствуя адсорбции вируса на клетках-мишенях. Напряженность противовирусного иммунитета оценивают преимущественно в серологических тестах -- по нарастанию титра специфических антител в парных сыворотках в процессе болезни. Иногда определяют концентрацию интерферона в сыворотке крови. Трансплантационным иммунитетом называют иммунную реакцию макроорганизма, направленную против пересаженной в него чужеродной ткани (трансплантата). Знание механизмов трансплантационного иммунитета необходимо для решения одной из важнейших проблем современной медицины -- пересадки органов и тканей. Многолетний опыт показал, что успех операции по пересадке чужеродных органов и тканей в подавляющем большинстве случаев зависит от иммунологической совместимости тканей донора и реципиента. Иммунная реакция на чужеродные клетки и ткани обусловлена тем, что в их составе содержатся генетически чужеродные для организма антигены. Эти антигены получили название трансплантационных, или антигенов гистосовместимости. Комплекс антигенов гистосовместимости наиболее полно представлен на цитоплазматической мембране лейкоцитов крови -- в системе HLA.
Реакция отторжения не возникает в случае полной совместимости донора и реципиента по антигенам гистосовместимости -- такое возможно лишь у однояйцовых близнецов. Выраженность реакции отторжения во многом зависит от степени чуже-родности и объема трансплантируемого материала. При контакте с чужеродными трансплантационными антигенами организм реагирует факторами клеточного и гуморального звеньев иммунитета. Основным фактором трансплантационного иммунитета являются Г-киллеры. Эти клетки после сенсибилизации антигенами донора мигрируют в пересаженную ткань (трансплантат) и оказывают на нее цитолитическое действие. Механизм иммунного отторжения пересаженных клеток и тканей имеет две фазы. В первой фазе вокруг трансплантата и сосудов наблюдается скопление иммунокомпетентных клеток (лимфоидная инфильтрация), в том числе сенсибилизированных Т-киллеров.
Во второй фазе клетки трансплантата разрушаются Т-киллерами, возникают воспаление и тромбоз кровеносных сосудов, нарушается питание трансплантата и последний гибнет. Погибшие клетки утилизируются фагоцитами. Формируется клон Т-клеток иммунологической памяти. Повторная попытка пересадки тех же органов и тканей вызывает вторичный иммунный ответ, который протекает очень бурно и быстро заканчивается реакцией отторжения трансплантата. Этот феномен назван кризом отторжения.
Особенности иммунитета при вирусных инфекциях
О напряженности противовирусного иммунитета судят преимущественно по нарастанию титра специфических антител в сыворотке крови больного в динамике заболевания или после специфической вакцинации. Защитные механизмы специфического, противовирусного иммунитета обеспечиваются также клетками-эффекторами (Т-киллеры, NK-клетки и другие клетки, участвующие в АЗЦТ). Специфические антитела против различных вирусных антигенов нередко присутствуют в сыворотках здоровых людей, что объясняется всеобщей иммунизацией населения против ряда вирусных инфекций (полиомиелит, корь, грипп и др.), а также возможностью скрытого (латентного) течения некоторых из них (герпес, гепатит и др.).
Неспецифические факторы иммунитета
Они защищают организм человека от всех заболеваний и обусловлены врожденными свойствами организма, которые способствуют уничтожению самых различных микроорганизмов на поверхности тела и его полостях К. неспецифическим факторам иммунитета относят:
Тканевые (клеточные) факторы. Среди тканевых факторов важную роль выполняют:
Иммунологические барьеры, к которым относят защитные свойства кожи, слизистых и лимфоузлов. Кожа и слизистые являются механическим барьером, секрет потовых, сальных желез и секрет слизистых угнетают многие виды патогенных микроорганизмов. Лимфоузлы препятствуют распространению микроорганизмов в макроорганизме, являясь мощным естественным барьером
Фагоцитоз - процесс активного поглощения клетками макроорганизма попавших в него чужеродных веществ (в т.ч. микроорганизмов) с последующим их перевариваем с помощью внутриклеточных ферментов. Стадии фагоцитоза: 1) приближение фагоцита к объекту - положительный хемотаксис; 2) прилипание микроорганизма к фагоцитам - адгезия; 3) поглощение (инвагинация) микроорганизмов фагоцитами и образование фагосомы; 4) образование фаголизосомы, переваривание и гибель микроорганизма - киллинг-инактивация. Различают завершенный фагоцитоз (заканчивается полным разрушением и гибелью микроорганизма) и незавершенный (микроорганизмы внутри фагоцита не только не гибнут, но даже размножаются). Фагоцитарной активностью обладают микрофаги (это гранулярные лейкоциты - нейтрофилы, эозинофилы, базофилы) и макрофаги (подвижные - моноциты крови, фиксированные - гистиоциты, эндотелиальные и ретикулярные клетки внутренних органов и костного мозга)
Видовая реактивность клеток - отсутствие рецепторов на поверхности клеток делает невозможным адсорбцию и проникновение инфекционного агента или яда в клетку
Нормальные киллеры (клетки убийцы, NK) - это большие лимфоциты с относительно рыхлым почковидным крупным ядром и обильной цитоплазмой с содержанием большого количества цитотоксических веществ, узнающие с помощью специальных рецепторов клетки-мишени, инфицированные вирусами, онкогенные клетки и разрушающие их.
Гуморальные факторы неспецифической защиты. Многочисленны, содержатся в крови и других жидкостях организма человека, вырабатываются Т-лимфоцитами и макрофагами. К ним относят:
Комплемент - неспецифическая ферментная система крови, состоящая из 9 различных протеиновых фракций, адсорбирующихся в процессе каскадного присоединения на комплексе антиген + антитело и оказывающих лизирующее действие на связанные антителами клеточные антигены
Лизоцим - белок, содержащийся в слюне, крови, слезной и тканевой жидкости, активен в отношении грамположительных бактерий, т.к. нарушает синтез муреина в клеточной стенке.
в-лизины - освобождаются из лейкоцитов и более активны по отношению к грамотрицательным бактериям
лейкины - протеолитические ферменты, освобождающиеся при разрушении лейкоцитов и нарушающие целостность поверхностных белков микробных клеток
интерферон - б и в, продуцируются соответственно мононуклеарными фагоцитами и фибробластами и обладают противовирусной активностью
пропердин - комплекс белков, обладающих противовирусной, антибактериальной активностью в присутствии солей магния, вызывая лизис микроорганизмов и усиливая фагоцитарную реакцию и воспалительный процесс
эритрин - обладает ингибирующим действием на коринебактерии дифтерии и высвобождается при разрушении эритроцитов
нормальные антитела - обнаруживаются в крови новорожденных в очень низких титрах, обладают цитофильным действием, уровень их возрастает под действием микроорганизма как пускового сигнала. Образование нормальных антител генетически запрограммировано, они экспрессируются на поверхностных мембранах незрелых В-лимфоцитов в виде рецепторов
Факторы саморегуляции: В основе антиинфекционного иммунитета лежит также способность организма к саморегуляции. Эта форма иммунитета, прежде всего, проявляются повышением температуры тела, губительно сказывающейся на жизнедеятельности многих бактерий и, особенно, вирусов в результате непосредственного влияния или изменения рН и окислительно-восстановительного потенциала (rН2) пораженных тканей. Большое значение в обеспечении естественного иммунитета имеет усиление выделительных функций организма, выведение микроорганизмов и их токсинов с мочой, испражнениями, мокротой и другими экскретами.
Неспецифический иммунитет
Во главу своей системы И. И. Мечников ставил фагоцит, или клетку. Против такой трактовки яростно выступали сторонники “гуморального” иммунитета Э. Беринг, Р. Кох, П. Эрлих (Нобелевские премии 1901, 1905 и 1908 гг.). Латинское “гумор” или “юмор” означает жидкость, в данном случае имелась в виду кровь и лимфа. Все трое считали, что организм защищается от микробов с помощью особых веществ, плавающих в гуморах. Их назвали “а н т и т о к с и н ы” и “а н т и т е л а”.
Нужно отметить прозорливость членов Нобелевского комитета, которые еще в 1908 г. попытались примирить две противоборствующие теории иммунитета, наградив И. И. Мечникова и немца Пауля Эрлиха. Потом премии иммунологам посыпались как из рога изобилия (см. Приложение).
Ученик Мечникова бельгиец Ж. Борде открыл в крови особое вещество.Оно оказалось белком, который помогает антителам распознать антиген.
А н т и г е н а м и называют вещества, которые при попадании в организм стимулируют выработку а н т и т е л. В свою очередь, антитела представляют собой высокоспецифические белки. Связываясь с антигенами (например бактериальными токсинами), они нейтрализуют их, не давая разрушать клетки. А н т и т е л а синтезируются в организме лимфоцитами или клетками лимфы. Л и м ф о й греки называли чистую и прозрачную воду подземных ключей и источников. Лимфа, в отличие от крови, прозрачная желтоватая жидкость. Лимфоциты находятся не только в лимфе, но и в крови. Однако попадания антигена в кровь еще не достаточно для того, чтобы начался синтез антител. Необходимо, чтобы антиген был поглощен и переработан фагоцитом, или макрофагом. Таким образом, мечниковский макрофаг стоит в самом начале иммунного ответа организма. Схема этого ответа может выглядеть следующим образом:
Антиген - Макрофаг - ? - Лимфоцит - Антитела - Инфекционный агент
Можно сказать, что вокруг этой простенькой схемки вот уже столетие кипят страсти. Иммунология стала теорией медицины и важной биологической проблемой. Здесь завязываются молекулярная и клеточная биология, генетика, эволюция и многие другие дисциплины. Неудивительно, что именно иммунологи получили львиную долю биомедицинских Нобелевских премий.
Воспаление как механизм неспецифического иммунитета
Воспаление - реакция организма на чужеродные микроорганизмы и продукты тканевого распада. Это основной механизм е с т е с т в е н н о г о (врожденного, или неспецифического) иммунитета, равно как начальный и заключительный этапы иммунитета п р и о б р е т е н н о г о. Как и всякая защитная реакция, оно должно сочетать способность распознавать чужеродную для организма частицу с действенным способом ее обезвреживания и удаления из организма. Классический пример - воспаление, вызванное занозой, прошедшей под кожу и загрязненной бактериями.
В норме стенки кровеносных сосудов непроницаемы для компонентов крови - плазмы и форменных элементов (эритроцитов и лейкоцитов). Повышенная проницаемость для плазмы крови -следствие изменения стенки сосудов, образования "щелей" между плотно прилегающими друг к другу клетками эндотелия. В районе занозы наблюдается торможение движения эритроцитов и лейкоцитов (клеток белой крови), которые начинают как бы липнуть к стенкам капилляров, образуя “пробки”. Два типа лейкоцитов - моноциты и нейтрофилы - начинают активно “протискиваться” из крови в окружающую ткань между клетками эндотелия в районе формирующегося воспаления.
Моноциты и нейтрофилы предназначены для фагоцитоза - поглощения и разрушения посторонних частиц. Целенаправленное активное движение к очагу воспаления носит название х е м о т а к с и с а. Придя к месту воспаления, моноциты превращаются в макрофаги. Это клетки с тканевой локализацией, активно фагоцитирующие, с “липкой” поверхностью, подвижные, как бы ощупывающие все, что находится в ближайшем окружении. Нейтрофилы также приходят в очаг воспаления, и их фагоцитирующая активность возрастает. Фагоцитирующие клетки накапливаются, активно поглощают и разрушают (внутриклеточно) бактерии и обломки клеток.
Активизация трех главных систем, участвующих в воспалении, определяет состав и динамику “действующих лиц”. Они включают систему образования кининов, систему комплемента и систему активированных фагоцитирующих клеток.
Образование кининов
Каким образом защитные клетки, циркулирующие в крови или осевшие в органах и тканях иммунной системы, получают и воспринимают сигналы опасности микробной агрессии? Как обеспечивается строгая последовательность включения отдельных типов клеток в борьбе с инфекцией?
Среди продуктов, нарабатываемых макрофагами в очаге инфекции, есть особые молекулы, получившие название фактор некроза опухолей (ФНО). Название связано с цитотоксичностью этих молекул, то есть с их способностью убивать клетки-мишени, в том числе опухолевые клетки. Рецепторы для этого фактора обнаружены на поверхности всех ядерных клеток организма, он способен вмешиваться в самые разные процессы. ФНО имеет непосредственное отношение к мобилизации клеток макрофагов в очагах инфекции.
Роль Т-лимфоцитов в иммунном ответе
Хотя иммунный ответ запускает макрофаг, только лимфоциты имеют специальные рецепторы для распознавания чужеродных молекул “антигенов” и обеспечивают иммунный ответ. Одновременно два сигнала активации идут с поверхности Т-лимфоцитов к ядру: от антиген-распознающего рецептора и от рецептора, связавшего ИЛ-1. Под действием этого двойного сигнала в геноме Т-лимфоцитов активируются гены как самого ИЛ-2, так и гены рецепторов, специфичных для ИЛ-2. После этого продукт Т-лимфоцитов ИЛ-2 начинает воздействовать на клетки, в которых он и был синтезирован: в этих клетках активируется процесс деления. В результате усиливаются функции всей популяции Т-лимфоцитов, участвующих в специфическом иммунном ответе на данный антиген (рис.8).
Характер иммунного ответа зависит от присутствия определенных цитокинов в микроокружении Т-лимфоцитов в момент распознавания антигена и активации. Если в этот момент в окружающей среде преобладает интерлейкин-4, клетки Т-лимфоцитов превращаются в активированных Т-хелперов (помощников) и начинают синтезировать тот же ИЛ-4, а также ИЛ-5,6,7,10. Эти интерлейкины активируют через соответствующие рецепторы деление В-лимфоцитов, их созревание в плазматические клетки, а также начинающийся синтез специфических для данного антигена антител-иммуноглобулинов. Это объясняет, почему в данном случае Т-лимфоциты выступают в роли Т-хелперов, то есть помощников В-лимфоцитов в их основном деле - наработке запаса защитных молекул - антител (см. рис. 8).
Но и этим не исчерпываются возможные функции Т-лимфоцитов. Они держат весь иммунный ответ под контролем, не допуская чрезмерной активации отдельных иммунокомпетентных клеток, которая чревата осложнениями. Инструментами такого контроля служат цитокины, способные не только активировать (усиливать), но и подавлять (ингибировать) функции других клеток.
Между Т-лимфоцитами и макрофагами существует двухсторонняя связь. Первые получают от макрофагов сигнал активации в виде молекулы интерлейкина-1, для восприятия которого имеют на поверхности соответствующие рецепторы (рис. 9). От рецепторов идет сигнал активации генов Т-лимфоцитов, заведующих синтезом ИЛ-2 и гамма-интерферона. Рецепторы Т-лимфоцитов распознают ИЛ-2. После того, как последний садится на рецептор, от него поступает сигнал дальнейшей активации синтезов в клетках Т-лимфоцитов и начала деления клетки. Что касается гамма-интерферона, то эти молекулы направляются в виде ответного послания макрофагу, на поверхности которого их ждут соответствующие рецепторы. Гамма-интерферон не зря называют макрофаг-активирующим фактором. Связавшись со своим рецептором на внешней поверхности клетки-макрофага, он посылает к ядру этой клетки сигналы активации нескольких десятков генов, в том числе гена, ответственного за синтез интерлейкина-1. В результате Т - лимфоциты получают от активированного макрофага новую порцию активирующих их молекул ИЛ-1 (рис. 9).
Система образования кининов обнаруживает чужеродное тело по его отрицательно заряженной поверхности. На ней адсорбируется так называемый фактор Хагемана (ФХ) - один из начальных компонентов системы свертывания крови. Этот белок присутствует в крови и имеет сродство к отрицательно заряженным поверхностям. Поверхности же собственных клеток устроены так, что они не адсорбируют ФХ и не индуцируют тем самым дальнейшую цепь событий. Это самый простой и примитивный способ отличать “свое” от “не своего”, используемый организмом в естественном иммунитете. Вторая особенность системы образования кининов - ряд каскадных усилений начальной реакции, резко повышающих эффект первичных взаимодействий.
Таким образом, “точечная” начальная реакция на чужеродной поверхности порождает макроскопические, видимые простым глазом физиологические изменения в формирующемся очаге воспаления.
иммунитет вирусный инфекция клетка
Система комплемента и ее активация
Комплементом называются сложный комплекс белков (около 20), которые, так же как и белки, участвующих в процессе свертывания крови, фибринолиза и образования кининов, формирует каскадные системы, обнаруженные в плазме крови. Для этих систем характерно формирование быстрого, многократно усиленного ответа на первичный сигнал за счет каскадного процесса. В этом случае продукт одной реакции служит катализатором последующей.
Ряд компонентов системы комплемента обозначают символом “С” и цифрой. В наибольшей концентрации в сыворотке крови присутствует компонент С3 (1,2 мг/мл). Система комплемента представлена, главным образом, неактивными предшественниками протеаз, действующих на белки. Активация системы в е с т е с т в е н н о м, то есть врожденном, иммунитете начинается с его третьего компонента С3 (рис. 10).
Конечный компонент системы комплемента (С9) включается в комплекс, атакующий мембрану бактерий. Присоединяя к себе несколько таких же, как и он сам, молекул, он погружается в мембрану и полимеризуется в кольцо. Образуются поры, “продырявливающие” оболочку бактерии, что ведет к ее гибели. Таким образом система комплемента распознает чужеродную клетку и запускает цепную реакцию активации биологически активных белков. Это ведет к приобретению комплексом токсической активности и гибели бактериальной клетки.
Т у ч н ы е к л е т к и активно синтезируют и хранят большие запасы мощного медиатора воспаления - гистамина. Тучные клетки рассеяны повсеместно в соединительной ткани и особенно вдоль кровеносных сосудов.
Когда к ним присоединяются пептиды, тучные клетки секретируют гистамин в окружающую среду. Эндотелий капилляров под его воздействием выделяет сосудорасширяющие вещества, и поток крови через очаг воспаления существенно возрастает. Между клетками эндотелия образуются “щели”, плазма выходит из капилляров в зону воспаления, свертывается и изолирует тем самым распространение инфекции из очага. По градиенту концентрации гистамина фагоциты “поднимаются” к источнику воспаления. Гистамин действует активно и быстро, благодаря чему и является медиатором острой фазы воспаления.
Возвращаясь к комплементу, следует еще раз подчеркнуть многонаправленность его действия (токсичность для микроорганизмов, усиление фагоцитоза, генерация медиаторов воспаления) и каскадное усиление всех направлений его активности.
Система комплемента стоит в центре формирования воспаления и прямо ведет к его важнейшему компоненту - фагоцитозу.
Фагоцитоз
Громадная роль фагоцитоза не только во врожденном, но и в приобретенном иммунитете становится все более очевидной благодаря работам последнего десятилетия. Фагоцитоз начинается с накопления фагоцитов в очаге воспаления. Главную роль в этом процессе играют моноциты и нейтрофилы. Моноциты, придя в очаг воспаления, превращаются в макрофаги - тканевые фагоцитирующие клетки. Фагоциты, взаимодействуя с бактериями, активируются, их мембрана становится “липкой”, в цитоплазме накапливаются гранулы, наполненные мощными протеазами. Возрастают поглощение кислорода и генерация активных форм кислорода (кислородный взрыв), включая перекиси водорода и гипохлорита, а также окись азота
В дополнение к перечисленным признакам активации, макрофаги начинают выделять в среду мощные медиаторы воспаления, среди которых особой активностью отличаются фактор некроза опухолей (ФНО), гамма-интерферон (Int-y) и интерлейкин-8 (ИЛ-8). Все они являются биологически активными пептидами.
Какова же их роль в воспалении? Начнем с ФНО. Этот небольшой белок, синтезируемый и секретируемый макрофагами, обладает множественной активностью. Он активирует сами же макрофаги и нейтрофилы, а также индуцирует синтез и появление на мембране клеток сосудистого эндотелия особых белков, специфически взаимодействующих с клеточной поверхностью моноцитов и нейтрофилов. Поверхность эндотелия благодаря этому становится “липкой” для этих клеток.
ИЛ-8 вызывает появление в клетках эндотелия рецепторов, реагирующих с моноцитами и нейтрофилами с высоким сродством, так что эти клетки останавливаются в капиллярах в районе воспаления. Именно IL-8, наряду с другими факторами воспаления, стимулирует моноциты и нейтрофилы к миграции по его градиенту в очаг воспаления. Фагоциты имеют рецепторы к интерлекину-8, которые “чувствуют” разницу в его концентрации и направляют свое движение по оси максимального отличия
Гамма-интерферон также “многоцелевой” медиатор воспаления. Одна из главных его функций - аутоактивация макрофагов и активация нейтрофилов. В этих клетках резко усиливается синтез протеаз, которые накапливаются в специальных цитоплазматических органеллах - лизосомах. В них происходит “кислородный взрыв” - продукция активных форм кислорода и окиси азота, высокотоксичных для микроорганизмов. Поверхность фагоцитов становится “липкой”, т.к. количество различных рецепторов на ней резко увеличивается, как увеличивается и “ощупывающая” подвижность цитоплазмы этих клеток. Когда такая клетка встречается с бактерией, то она "прилипает" к поверхности фагоцита, обволакивается его псевдоподиями и оказывается внутри клетки (где и разрушается протеазами).
Так заканчивается цикл “воспаление - фагоцитоз”. Мы видим, насколько мощный и многогранный защитный механизм действует в этом случае. Однако его слабость в его однообразии, как бы в монотонности механизма, срабатывающего одинаково при встрече с различными врагами. Естественно, что многообразные и многочисленные популяции микроорганизмов вырабатывают обходные пути для проникновения в организм.
Специфический иммунитет
А. Гуморальный и клеточный иммунитет
В зависимости от функций лимфоцитов, специфический иммунитет принято делить также на г у м о р а л ь н ы й и к л е т о ч н ы й. В-лимфоциты в данном случае ответственны за гуморальный, а Т-лимфоциты - за клеточный иммунитет. Гуморальный иммунитет назван так потому, что его иммуноциты (В-клетки) вырабатывают антитела, способные отделяться от клеточной поверхности. Продвигаясь по кровяному или лимфатическому руслу - гумору (от. лат. humor - жидкость), антитела поражают чужеродные тела на любой дистанции от лимфоцита. К л е т о ч н ы м иммунитет именуют потому, что Т-лимфоциты (преимущественно Т-киллеры) вырабатывают рецепторы, жестко фиксированные на клеточной мембране, и служат Т-киллерам эффективным оружием для поражения чужеродных клеток при непосредственном контакте с ними.
На периферии зрелые Т- и В-клетки располагаются в одних и тех же лимфоидных органах - частично изолированно, частично в смеси. Но что касается Т-лимфоцитов, то их пребывание в органах непродолжительно. “Вечные странники” постоянно в движении. Срок их жизни (месяцы и годы) способствует им в этом. Т-лимфоциты многократно покидают лимфоидные органы, попадая сначала в лимфу, затем в кровь, а из крови снова возвращаются в органы. За свою жизнь лимфоцит проходит поразительно большие расстояния - от 100 до 1 млн. км. Благодаря циркуляции лимфоциты удивительно быстро появляются в “горячих точках”. Без такой способности лимфоцитов были бы невозможны своевременное их развитие, взаимодействие и эффективное участие в и м м у н н о м о т в е т е при вторжении чужеродных молекул и клеток.
Полноценное развитие г у м о р а л ь н о г о иммунного ответа требует не двух, а по крайней мере трех типов клеток. Функция каждого клеточного типа в антителопродукции строго предопределена. Макрофаги и другие фагоцитирующие клетки поглощают, перерабатывают и экспрессируют антиген в иммуногенной, доступной для Т- и В- лимфоцитов форме. Т-хелперы после распознавания антигена начинают продукцию цитокинов, обеспечивающих помощь В-клеткам. Эти последние клетки, получив специфический стимул от антигена и неспецифический от Т-клеток, начинают продукцию антител (рис. 13).
При формировании к л е т о ч н о г о типа иммунного ответа также необходима кооперация различных типов клеток.
Б. Характерные черты специфического иммунитета
Рассмотрим реакцию с п е ц и ф и ч е с к г о иммунитета на двух внешне совершенно непохожих моделях - выведении чужеродного белка и отторжении чужеродной ткани.
Чужеродный белок не вызывает ни воспаления, ни фагоцитоза. Он беспрепятственно минует первую линию защиты и непосредственно встречается со второй. Организм отличает “свое” от “не своего”, это первая особенность реакции специфического иммунитета. Так называемая иммунологическая память - вторая характерная черта реакции специфического иммунитета. Память специфична, запоминается контакт лишь с определенным белком. Специфичность запоминания очень высока, и это третья особенность реакции приобретенного иммунитета.
Иммунный ответ на чужеродную макромолекулу можно избирательно подавить, если ввести ее в развивающийся организм внутриутробно или в первые часы после рождения. Способность отличать введенный чужеродный белок от собственного у такого животного утрачивается после рождения. Подавление реакции строго специфично - оно распространяется только на белок, введенный в процессе развития. Это явление носит название т о л е р а н т н о с т и (терпимости). Оно составляет четвертую неотъемлемую особенность элементарной реакции п р и о б р е т е н н о г о, или с п е ц и ф и ч е с к о г о, иммунитета. Четыре признака реакции неразделимы, они всегда вместе.
Приобретенный иммунитет широко используется для вакцинации. Введение ослабленных или убитых микробов (или выделенных из них макромолекул) вызывает иммунологическую реакцию. Вакцинация является основным способом предупреждения таких страшных заболеваний как оспа, туберкулез, полиомиелит, сибирская язва и многих других. Приобретенный иммунитет составляет основное препятствие для пересадок органов (сердце, почки, печень) и ткани (кожа) от одного человека к другому. Для преодоления этого барьера несовместимости пользуются препаратами, подавляющими иммунную систему.
В. Антитела и антиген-распознающие рецепторы лимфоцитов
Вещество, способное вызывать реакцию приобретенного иммунитета, носит название а н т и г е н а. Антигеном может быть не всякое вещество. Оно должно быть чужеродным, макромолекулярным и иметь устойчивую химическую структуру. К типичным антигенам относятся белки и полисахариды.
Когда свободный антиген циркулирует в крови, в ответ на него появляются защитные белки - антитела, которые специфически распознают его и образуют с ним комплекс. Антитела обезвреживают антиген, если он токсичен (бактериальный токсин, змеиный яд или болезнетворный вирус), и способствуют выведению его из организма.
При отторжении трансплантата главная роль принадлежит особой разновидности лимфоцитов Т - к и л л е р а м, “убийцам”. Эти лимфоциты несут на своей наружной мембране антителоподобные рецепторы - рецепторы Т-клеток (РТК). РТК специфически распознают антиген, находящийся на мембране чужеродных клеток, и обеспечивают прикрепление Т-киллера к клетке-мишени. Это необходимо для осуществления киллером своей смертоносной функции. Прикрепившись, они выделяют в просвет между собой и мишенью особый белок, “продырявливающий” мембрану клетки-мишени, в результате чего клетка гибнет. Затем они открепляются от мишени и переходят на другую клетку, и так несколько раз. При повторной встрече организма с антигеном образуется больше антител и киллеров, они появляются в более короткие сроки, чем при первой встрече. В этом и проявляется иммулогическая память, и на этом основан эффект вакцинации
Как устроены антитела и рецепторы лимфоцитов? Очевидно, что их структура должна быть необычной, так как они строго специфически распознают громадное множество различных антигенов - любой чужеродный белок, полисахарид или синтетическую молекулу, вообще в природе не встречающиеся. При этом антитела и Т-лимфоциты, как правило, не реагируют с белками и клетками собственного организма. Как природа решает эту задачу?
Принцип строения антитела и РТК представлен на рис. 15. Часть молекулы антитела, которая связывается с антигеном, очень изменчива, поэтому ее называют в а - р и а б е л ь но й (V). V-часть концентрируется на самом кончике антитела, поэтому защитную молекулу можно сравнить с пинцетом, ухватывающим с помощью острых концов мельчайшие детали самого замысловатого часового механизма. Активный центр распознает в молекуле антигена небольшие участки, состоящие из 4 - 8 аминокислот. Эти участки антигена подходят к структуре активного центра, “как ключ к замку”.
Разнообразие активных центров очень велико, не менее 107 -10 8. Этого вполне достаточно для специфического распознавания любого произвольно взятого антигена антителами и РТК (а точнее, их активными центрами).
Следует подчеркнуть, что весь репертуар антител или РТК. он возникает до встречи с антигеном и независимо от антигена, а большая часть антител не потребуется в течение всей жизни особи.
Г. Клеточные механизмы иммунитета
Как сказано, в организме предсуществуют антитела и РТК к любому произвольно взятому антигену. Эти антитела и РТК присутствуют на поверхности лимфоцитов, образуя там антигенраспознающие рецепторы. Чрезвычайно важно, что один лимфоцит может синтезировать антитела (или РТК) только одной специфичности, не отличающиеся друг от друга по структуре активного центра. Это формулируется как принцип “один лимфоцит - одно и антитело”.
Каким же образом антиген, попадая в организм, вызывает усиленный синтез именно тех антител, которые специфично реагируют только с ними? Ответ на этот вопрос дала теория селекции клонов австралийского исследователя Ф.М. Бернета. Согласно этой теории, одна клетка синтезирует лишь один тип антител, которые локализуются на ее поверхности. Репертуар антител формируется до и независимо от встречи с антигеном. Роль антигена заключатся лишь в том, чтобы найти клетку, несущую на своей мембране антитело, реагирующее именно с ним, и активировать эту клетку. Активированный лимфоцит вступает в деление и дифференцировку. В результате из одной клетки возникает 500 - 1000 генетически идентичных клеток (клон). Клон синтезирует один и тот же тип антител, способных специфически распознавать антиген и соединяться с ним (рис. 16). В этом и заключается суть иммунного ответа: селекции нужных клонов и их стимуляции к делению.
В основе образования лимфоцитов-киллеров тот же принцип: селекция антигенов Т-лимфоцита, несущего на своей поверхности РТК нужной специфичности, и стимуляция его деления и дифференцировки. В результате образуется клон однотипных Т-киллеров. Они несут на своей поверхности большое количество РТК. Последние взаимодействуют с антигеном, входящим в состав чужеродной клетки, и способных убивают эти клетки.
Киллер ничего не может сделать с растворимым антигеном - ни обезвредить его, ни удалить из организма. Но лимфоцит-киллер очень активно убивает клетки, содержащие чужеродный антиген. Поэтому он проходит мимо растворимого антигена, но не пропускает антиген, находящийся на поверхности “чужой” клетки.
Детальное изучение реакции иммунитета показало, что для образования клона клеток, продуцирующих антитела, или клона Т-киллеров необходимо участие специальных лимфоцитов-помощников (Т-хэлперов). Сами по себе они не способны ни вырабатывать антитела, ни убивать клетки-мишени. Но, распознавая чужеродный антиген, они реагируют на него выработкой ростовых и дифференцированных факторов. Данные факторы необходимы для размножения и созревания антителообразующих и киллерных лимфоцитов. В этой связи интересно вспомнить о вирусе СПИДа, вызывающем сильное поражение иммунной системы. Вирус ВИЧ поражает именно Т-хэлперы, делая иммунную систему не способной ни к выработке антител, ни к образованию Т-киллеров.
Д. Эффекторные механизмы иммунитета
Как антитела или Т-киллеры удаляют из организма чужеродные вещества или клетки? В случае киллеров РТК выполняют лишь функцию “наводчика” - они распознают соответствующие мишени и прикрепляют к ним клетку-убийцу. Так распознаются клетки, зараженные вирусом. Сам по себе РТК не опасен для клетки-мишени, но “идущие за ним” Т-клетки представляют огромный разрушительный потенциал. В случае антител мы встречаемся со сходной ситуацией. Сами по себе антитела безвредны для клеток, несущих антиген, но при встрече с антигенами, циркулирующими или входящими в состав клеточной стенки микроорганизма, к антителам подключается с и с т е м а к о м п л е м е н т а. Она резко усиливает действие антител. Комплемент сообщает образующемуся комплексу антиген - антитело биологическую активность: токсичность, сродство с фагоцитирующим клеткам и способность вызывать воспаление.
Первый компонент этой системы (С3) распознает комплекс антиген - антитело. Распознавание ведет к появлению у него ферментативной активности к последующему компоненту. Последовательная активация всех компонентов системы комплемента имеет ряд последствий. Во-первых, происходит каскадное усиление реакции. При этом продуктов реакции образуется несравнимо больше, чем исходных реагирующие веществ. Во-вторых, на поверхности бактерии фиксируются компоненты (С9) комплемента, резко усиливающих фагоцитоз этих клеток. В-третьих, при ферментативном расщеплении белков системы комплемента образуются фрагменты, обладающие мощной воспалительной активностью. И, наконец, при включении в комплекс антиген-антитело последнего компонента комплемента этот комплекс приобретает способность “продырявливать” клеточную мембрану и тем самым убивать чужеродные клетки. Таким образом, система комплемента - важнейшее звено в защитных реакциях организма.
Однако комплемент активируется любым комплексом антиген-антитело, вредным или безвредным для организма. Воспалительная реакция на безвредные антигены, регулярно попадающие в организм, может вести к аллергическим, то есть извращенным, реакциям иммунитета. Аллергия развивается при повторном попадания антигена в организм. Например, при повторном введении антитоксичных сывороток, или у мукомолов на белки муки, или при многократной инъекции фармацевтических препаратов (в частности, некоторых антибиотиков). Борьба с аллергическими болезнями состоит в подавлении либо самой реакции иммунитета, либо в нейтрализации образующихся при аллергии веществ, вызывающих воспаление.
Размещено на Allbest.ru
...Обзор механизмов лимфоидного аппарата адаптивного иммунитета. Система образования кининов. Рецепторы клеток врожденной иммунной системы. Характеристика сигналов и их реализации. Особенности взаимодействия плазменных белков, их участие в иммунных реакциях.
курсовая работа , добавлен 02.03.2013
Исследование иммунной системы человека. Изучение особенностей формирования неспецифического иммунитета. Анализ естественной, врожденной и приобретенной форм иммунитета. Описания функций клеток памяти и эффекторов, системы комплемента, структуры антигена.
презентация , добавлен 13.12.2012
Иммунитет – способ защиты организма от болезнетворных микроорганизмов за счет выработки антител. Обзор схемы клеточного и гуморального иммунитета. Нарушения фагоцитарной системы. Методы оценки иммунитета. Реакция иммунного гемолиза и цитотоксический тест.
презентация , добавлен 11.11.2014
Понятие и виды иммунитета, назначение иммунной системы. Факторы и признаки ослабления иммунитета, методы его повышения. Механизм действия иммунитета: макрофаги, Т-хэлперы, В-лимфоциты, выработка иммуноглобулинов (антител), Т-супрессоры, клетки-киллеры.
реферат , добавлен 09.02.2009
Формы, механизмы, органы, регуляция иммунитета. Субпопуляции Т-лимфоцитов, их функции. История открытия регуляторных Т-клеток. Эффективность микробиологической диагностики. Иммунная регуляторная система. Будущее трансплантологии, технические трудности.
контрольная работа , добавлен 11.05.2016
Система иммунитета организма и ее функции. Виды клеток иммунной системы (лимфоциты, фагоциты, гранулярные лейкоциты, тучные клетки, некоторые эпителиальные и ретикулярные клетки). Селезенка как фильтр крови. Клетки-убийцы как мощное оружие иммунитета.
презентация , добавлен 13.12.2015
Исследование роли микроорганизмов в процессах аммонификации, нитрофикации, денитрификации. Виды иммунитета - реакции организма, направленной на его защиту от внедрения чужеродного материала. Разложение пектиновых веществ. Анаэробное брожение клетчатки.
контрольная работа , добавлен 19.05.2012
Жизненный и творческий путь Ильи Ильича Мечникова – выдающегося русского ученого-биолога. Вклад Мечникова в развитие иммунологии. Фагоцитарная теория иммунитета. Развитие идей И.И. Мечникова в России и за рубежом, их практическое воплощение в жизнь.
реферат , добавлен 25.05.2017
Специфические факторы противовирусного иммунитета и синтез антител к определенному антигену. Клетки памяти и выдача иммунного ответа в форме биосинтеза антител. Распространение инфекционного бронхита птиц и ящера. Культивирование вирусов в клетках.
контрольная работа , добавлен 17.11.2010
Общая характеристика B-лимфоцитов. Характеристика субпопуляций, рецепторы и маркеры В-лимфоцитов. Антигенраспознающие рецепторы B-клеток: общая характеристика. Субпопуляции В-лимфоцитов, распознание антигенов рецепторами иммуноглобулиновой природы.
Иммунная система настолько качественно и точно работает, что для каждого вида инфекционного заболевания она использует свои механизмы и реакции. У каждого способа иммунного ответа существуют свои специфические черты. Особенности противовирусного иммунитета, антибактериального, противогрибкового состоят из принципов работы общего и местного иммунитетов.
Иммунитет - это невосприимчивость и устойчивость к патогенным и генетически чужеродным объектам.
Органы иммунной системы центральные, вырабатывающие защитные клетки, и периферические, контролирующие жидкие среды организма, кожные и слизистые покровы, а так же осуществляют контроль тканевых мигрирующих клеток.
В основе деятельности иммунитета лежит активное скооперированное воздействие необходимых частей системы на внедрение антигена, распознание чужеродного гена и уничтожение, то есть соответствующий ответ.
Иммунный ответ напрямую зависит от типа антигена, быстроты наступления иммунной реакции, внешних условий.
Иммунологический ответ бывает:
Самым распространенным возбудителем инфекционных заболеваний являются вирусные молекулы.
Болезни человека вызывают вирусы с суперкаспидами с отростками в виде шипов или пепломеров. У данного типа оболочка имеет дополнительный слой, состоящий из белковых вирусных частиц и липидов зараженных клеток.
Вирусное влияние наблюдается во всех формах жизни: от бактерий до человека.

Вирусная молекула начинает свой путь заражения с присоединения к специфическому рецептору, находящемуся на мембране здоровой клетки. После прикрепления, вирус начинает процессы:
Далее происходит разрушение вирусной оболочки, в следствие, высвобождается ген. Освободившаяся генетическая информация провоцирует синтезирование зараженной клеткой вирусных белковых компонентов, происходит удвоение и размножение вируса, его выход из погибшей клетки и распространение по другим соответствующим клеточным мембранам.
Для внедрения вирусной молекулы в организм главным препятствием являются барьерные механизмы местного и общего иммунитетов.
Особенности противовирусного иммунитета состоит в том, что он действует в двух направлениях:
Главной чертой противовирусного иммунного ответа является наличие специфических антител, которые в первом случае передаются по наследству, а во втором вырабатываются в момент возникновения вирусного заболевания.

Особенности врожденного антивирус иммунитета:

Можно говорить о том, что главным механизмом врожденного противовирусного иммунного сопротивления является интерферон. Находясь в достаточном количестве, стимулирует невосприимчивость иммунитета на клеточном уровне. Различают интерферон по видам:
Данное вещество не способствуют блокированию вирусных молекул, их функциональное действие направлено на изменение метаболических процессов клеток, вызывая устойчивость к вирусу.
Интерфероновые вещества продуцируются иммунными клетками под действием вирусных антигенов или после применения интерферон-содержащих препаратов.
Особенности противовирусного иммунитета приобретенного типа заключаются в том, что резистентность приобретается при перенесении заболевания или путем вакцинирования, и зависит от наличия специфических иммунных механизмов G, М, антител секреторного типа, иммунных Т-киллеров.
Находясь в крови даже в небольших количествах специфические антитела имеют способность осаждать и предотвращать развитие вируса, блокируя зараженные клетки. При наличии большого количества антигена происходит блокада внеклеточного вирусного объекта, находящегося в крови. Антитела способствует защите организма от вторичного заражения вирусом определенного типа.
Заключенные в блокаду вирусы уничтожаются лизисом системы комплимента.
Для нейтрализации вирусного заболевания противовирусным приобретенным иммунитетом используются вирусспецифические Т-киллеры, приводящие к развитию безвредной инфекции, но для иммунного действия такого рода должно быть достаточное количество.

В большинстве случаев иммунная реакция на проникновение вирусной инфекции складывается из:
Схема защитной реакции:
Проникновение вируса в организм препятствуют постоянное функционирование и слизистых. Если по каким-либо причинам местные функции иммунитета не справились со своей функцией, вирус проникает в организм начинает заражать здоровые клетки. Здесь на пути заболевания встают иммуннокомпетентные клетки иммунитета:
Специфические антитела блокируют зараженную клетку и белок вирусной оболочки, чем провоцируют их уничтожение лимфоцитами типа Т имеющими цитотоксическую функцию.

Какой вирус нарушает работу иммунной системы человека - это все вирусы, которые способны:
Нарушение специфической и неспецифической резистентности связано с:
Но неузнаваемые антителами вирусы могут без всяких препятствий распространиться по определенным здоровым клеткам, развивая вирусную болезнь. А вследствие эпидемии и пандемии.

Вирусные заболевания, к примеру , сопровождаются выраженными клиническими симптомами: температурой, насморком, кашлем, головной болью, общей слабостью, а так же характерными признаками: высыпаниями, расстройством пищеварения и другие. Для постановки полноценного диагноза проводится внешний осмотр, пальпация, внешние манипуляции, берутся соответствующие анализы. И назначается лечение:
Для защиты от вируса или для более легкого перенесения заболевания используются меры профилактики:
Имеющиеся способы противовирусной вакцинации.
Ранняя стадия инфекции, как правило, состоит в противоборстве вируса с защитными системами организма-хозяина. Самый первый защитный барьер - кожные покровы и слизистые оболочки организма. В случае нарушения их целостности в действие вступают механизмы экстренной неспецифической защиты (факторы врожденного иммунитета). Среди них особо выделяют противовирусную активность интерферона, ЕК-клетки (естественные киллеры) и макрофаги.
Противовирусное действие интерферона . Инфицирование клетки вирусом вызывает синтез интерферона. Под его действием активируются защитные механизмы соседних клеток, обеспечивая их устойчивость к вирусной инфекции. Интерферон индуцирует синтез двух ферментов: протеинкеназы, что ведет к подавлению синтеза вирусных белков, и 2", 5"-олигоаденилатсинтетазы, активирующей эндонуклеазу, которая разрушает вирусную и-РНК. Кроме того, интерферон сильно активирует макрофаги и ЕК-клетки.
Противовирусное действие ЕК-клеток и макрофагов . Активные ЕК-клетки появляются уже через двое суток после заражения организма-хозяина вирусом. ЕК-клетки и макрофаги уничтожают зараженные клетки. Главным образом ЕК-клетки осуществляют реакцию антителозависимой клеточной цитотоксичности (АЗКЦ).
Если вирусу удается преодолеть барьеры врожденной защиты, он вызывает развитие специфического иммунного ответа с появлением Т-киллеров, Т-хелперов и противовирусных антител. Главную роль в иммунном ответе отводят антителам и Т-киллерам. Основные механизмы противовирусного иммунитета сводятся к блокаде распространения вирусных частиц и уничтожению зараженных вирусом клеток, т.е. клеток, которые фактически являются «фабриками» по производству новых вирусов.
Распространение вируса в организме блокируют в основном антитела. В процессе развития специфического иммунитета синтезируются антитела к большинству антигенов вируса. Однако считается, что вирусную инфекцию сдерживают в основном антитела к поверхностным гликопротеинам. Эти антигены, часто называемые протективными, локализованы на поверхности вирионов или экспрессируются на мембране зараженной вирусом клетки. Механизмы гуморального противовирусного иммунитета могут быть различными. Способ устранения инфекционности вирусных частиц зависит от их локализации - внеклеточной или внутриклеточной.
Антитела, адсорбируясь на поверхности вирионов, блокируют его жизненно важные функции. Прежде всего, это блокада прикрепления к клетке хозяина, проникновения в нее, раздевание вируса. Адсорбция антител на белках капсида не позволяет некоторым вирусам (вирусу чумы плотоядных, кори и др.) проникать из клетки в клетку путем их слияния. Кроме того, считают, что антитела, активируя систему комплемента, вызывают повреждения оболочки некоторых вирусов и блокируют клеточные рецепторы для вирусов. Однако в настоящее время этот процесс не считают существенным в противовирусной защите.
Действие антител, помимо нейтрализации внеклеточных вирусов, состоит в том, что они вызывают разрушение инфицированных вирусами клеток, активируя систему комплемента. Второй механизм действия антител на внутриклеточный вирус - это реакция антителозависимой клеточной цитотоксичности, осуществляемая ЕК-клетками. Антитела, фиксированные на мембране пораженной вирусом клетки, контактируют с ЕК-клетками (через Fc-фрагмент IgG), которые убивают зараженные клетки с помощью перфоринов и гранзимов.
В иммунитете к вирусным инфекциям Т-клетки выполняют разнообразные функции. Т-хелперы играют важную роль в образовании антител в ответ на антигены, кроме того, эти клетки помогают в индукции Т-киллеров, а также в привлечении макрофагов и Е-клеток в очаг вирусной инфекции и в их активации. Т-киллеры осуществляют противовирусный иммунологический надзор, и действуют они весьма эффективно и избирательно, разрушая инфицированные вирусами клетки с помощью перфоринов и гранзимов. Проникнув внутрь клетки-мишени, гранзимы через каскад реакций активируют эндонуклеазы. Этот фермент способствует разрыву цепей ДНК и развитию апоптоза (программированная гибель клеток).
Механизмы «ухода» вирусов от иммунного надзора организма хозяина. Вирусы обладают разнообразными свойствами защиты от распознавания их антителами: наиболее эффективно этому служит смена антигенов: в вирусных белках происходит изменение иммунодоминатных областей. Антигенная изменчивость наблюдается у вирусов иммунодефицита человека и у вирусов гриппа. Так, у вируса гриппа она называется антигенным «дрейфом» (постепенные изменения) и «шифтом» (резкие изменения). Гуморальный иммунитет к этим вирусным инфекциям сохраняется лишь до появления нового сероварианта возбудителя, что не позволяет рассчитывать на долговременный эффект вакцинации; антитела могут удалять вирусные антигены с плазматической мембраны клетки путем кеппинга (агрегации молекул клеточной поверхности). Так, герпесвирусы кодируют гликопротеины, связывающие антитела через Fc-фрагмент, при этом нарушается активация комплемента и блокируется действие противовирусных антител; ряд вирусов (цитомегало-, аденовирусы и др.) индуцируют выработку белков, подавляющих экспрессию молекул ГКГС класса на мембране пораженных клеток. Это дает вирусу преимущество, помогая избежать распознавание эти отдельные вирусы (герпесвирусы) обладают генами белков, гомологичных цитокиновым рецепторам. В результате эти «растворимые» рецепторы как «ловушки» связывают цитокины и нейтрализуют их действия; некоторые вирусы (вирус Эпштейна - Барра, аденовирусы) способны противодействовать эффекту интерферонов - они продуцируют короткие отрезки РНК, которые каким-то образом подавляют активацию протеинкиназы; многие вирусы способны вызывать у макрофагов выработку супрессирующих цитокинов, подавляющих развитие иммунного ответа.
Ответ
Репродукция вирусов человека: этапы репродукции, особенности репродукции ДНК-содержащих вирусов. Приведите примеры таких вирусов. Эффект интерференции между вирусами. Типы взаимодействия вирусов с клеткой: продуктивный, абортивный, интегративный. Типы вирусных инфекций: литическая, персистирующая, латентная, иннапаратная, медленная вирусная инфекция, трансформация клетки.
Диагностические сыворотки, их подразделение, получение и практическое применение. Моноклональные антитела. Гибридомы, их использование для получения моноклональных антител.
Анатоксины: свойства, принцип получения, единицы измерения. Ассоциированные вакцины, их свойства, примеры. Охарактеризуйте иммунитет, формируемый в результате введения ассоциированных вакцин.
Серологические реакции, используемые в инфекционной иммунологии(см.20).Реакция иммунофлюоресценции. Ингредиенты,механизм прямой и непрямой РИФ.Значение для экспресс диагностики инфекционных заболеваний.
Иммунофлюоресцентный метод (РИФ, реакция иммунофлюоресценции, реакция Кунса) - метод выявления специфических АГ с помощью АТ, конъюгированных с флюорохромом. Обладает высокой чувствительностью и специфичностью.
Применяется для экспресс-диагностики инфекционных заболеваний (идентификация возбудителя в исследуемом материале), а также для определения АТ и поверхностных рецепторов и маркеров лейкоцитов (иммунофенотипирование) и др. клеток.
Обнаружение бактериальных и вирусных антигенов в инфекционных материалах, тканях животных и культурах клеток при помощи флюоресцирующих антител (сывороток) получило широкое применение в диагностической практике. Приготовление флюоресцирующих сывороток основано на способности некоторых флюорохромов (например, изотиоцианата флюоресцеина) вступать в химическую связь с сывороточными белками, не нарушая их иммунологической специфичности.
Различают три разновидности метода: прямой, непрямой, с комплементом. Прямой метод РИФ
основан на том, что антигены тканей или микробы, обработанные иммунными сыворотками с антителами, меченными флюорохромами, способны светиться в УФ-лучах люминесцентного микроскопа. Бактерии в мазке, обработанные такой люминесцирующей сывороткой, светятся по периферии клетки в виде каймы зеленого цвета.
Непрямой метод РИФ
заключается в выявлении комплекса антиген - антитело с помощью антиглобулиновой (против антитела) сыворотки, меченной флюорохромом. Для этого мазки из взвеси микробов обрабатывают антителами антимикробной кроличьей диагностической сыворотки. Затем антитела, не связавшиеся антигенами микробов, отмывают, а оставшиеся на микробах антитела выявляют, обрабатывая мазок антиглобулиновой (антикроличьей) сывороткой, меченной флюорохромами. В результате образуется комплекс микроб + антимикробные кроличьи антитела + антикроличьи антитела, меченные флюорохромом. Этот комплекс наблюдают в люминесцентном микроскопе, как и при прямом методе.
Механизм. На предметном стекле готовят мазок из исследуемого материала, фиксируют на пламени и обрабатывают иммунной кроличьей сывороткой, содержащей антитела против антигенов возбудителя. Для образования комплекса антиген - антитело препарат помещают во влажную камеру и инкубируют при 37 °С в течение 15 мин, после чего тщательно промывают изотоническим раствором хлорида натрия для удаления не связавшихся с антигеном антител. Затем на препарат наносят флюоресцирующую антиглобулиновую сыворотку против глобулинов кролика, выдерживают в течение 15 мин при 37 °С, а затем препарат тщательно промывают изотоническим раствором хлорида натрия. В результате связывания флюоресцирующей антиглобулиновой сыворотки с фиксированными на антигене специфическими антителами образуются светящиеся комплексы антиген - антитело, которые обнаруживаются при люминесцентной микроскопии.
25.Серологические реакции,применяемые в инфекционной иммунологии.(см.20)Иммуноблотинг, радиоиммунологический анализ:спецефичность,чувствительность,механизмы реакции.Практическое использование.
Иммуноблоттинг - высокочувствительный метод выявления белков, основанный на сочетании электрофореза и ИФА или РИА. Иммуноблоттинг используют как диагностический метод при ВИЧ-инфекции и др.
Антигены возбудителя разделяют с помощью электрофореза в полиакриламидном геле, затем переносят их из геля на активированную бумагуили нитроцеллюлозную мембрану и проявляют с помощью ИФА. Фирмы выпускают такие полоски с «блотами» антигенов. На эти полоски наносят сыворотку больного. Затем, после инкубации, отмывают от несвязавшихся антител больного и наносят сыворотку против иммуноглобулинов человека, меченную ферментом. Образовавшийся на полоске комплекс [антиген + антитело больного + антитело против Ig человека] выявляют добавлением хромогенного субстрата, изменяющего окраску под действием фермента.
С помощью РИА в биологических жидкостях определяют концентрации гормонов, факторов роста, ферментов, аутоантител, маркеров злокачественных новообразований и других веществ (например, лекарственных средств и наркотиков).
В основе РИА лежит феномен конкуренции: связывание антител с антигеном, меченным радиоактивным изотопом, подавляется в присутствии немеченого антигена.
Методика РИА проста и включает следующие основные этапы:
1. К антителам добавляют меченый антиген и пробу (содержащую неизвестное количество немеченого антигена). Концентрацию антител в реакционной смеси подбирают так, чтобы число мест связывания было намного меньше общего числа антигенов. Концентрация меченого антигена должна превышать максимально возможную концентрацию антигена в пробе.
2. Реакционную смесь инкубируют при определенной температуре заданное время. Меченный и немеченый антигены конкурентно связываются с антителами, при этом образуются иммунные комплексы, содержащие либо меченный, либо немеченый антиген. Таким образом, к концу инкубации в реакционной смеси присутствуют меченные и немеченые иммунные комплексы, а также свободные меченные и немеченые антигены. Количество меченных иммунных комплексов обратно пропорционально количеству немеченого антигена в пробе.
3. Чтобы оценить количество образовавшихся меченных иммунных комплексов, их отделяют от оставшегося несвязанным свободного меченого антигена.
4. Определяют концентрацию антигена в пробе по калибровочной кривой. Для ее построения используют несколько стандартных калибровочных растворов с известными концентрациями немеченого антигена.
26. Вакцины: их подразделение, основные требования, предъявляемые к вакцинам. Принципы получения инактивированных вакцин. Приведите примеры вакцин, применяемых для плановой вакцинации детей. Охарактеризуйте поствакцинальный иммунитет.
ОТВЕТ:
Вакцины - иммунобиологические препараты, изготовляемые из живых аттенуированных или инактивированных микроорганизмов, токсинов, микробных антигенов и используемые для создания специфического активного искусственного иммунитета.
В основном вакцины применяют с профилактической целью, значительно реже- с лечебной(при хронических затяжных инфекционных заболеваниях)
Подразделение вакцинных препаратов:
По составу на корпускулярные (живые и инактивированные), растворимые (химические и анатоксины) и генно-инженерные; по назначению на профилактические и лечебные.
Различаются несколько поколений вакцин:
-вакцины первого поколения - корпускулярные вакцины состоящие из целых микроорганизмов живых или убитых
-вакцины второго поколения- препараты состоящие из отдельных фракций возбудителя или продуктов их жизнедеятельности -химические вакцины и анатоксины
-вакцины третьего поколения -рекомбинатные вакцины, полученные генно-инженерными методами
Основные требования к вакцинных препаратам:
-высокая иммуногенность и создание достаточно стойкого иммунитета
-остаточная вирулентность для аттенуированных штампов и стабильность их свойств
-безвредность
-ареактивность (отсутствие выраденных побочных реакций)
-гипоаллергенность (минимальное сенсибилизирующее действие)
- отсутствие в препарате контаминирующиз микроорганизмов
-доступность стоимости производства
Инактивированные (убитые) корпускулярные клетки содержат микробные клетки или вирионы (корпускулярные бактериальные и цельновирионные вакцины). Для их приготовления используют вирулентные микроорганизмы, содержащие протективные антигены, активность которых должна сохраняться после воздействия инактивирующих факторов физической (нагревание, ультрафиолетовое излучение) или химической природы (спирт, фенол, ацетон, глутаровый альдегид, формальдегид) или комбинацией обоих факторов.
Инактивированные вакцины вызывают иммунный ответ гуморального типа, менее напряженной иммунитет с меньшей длительностью, чем живые вакцины, не индуцируют местный иммунитет, требуется их 2-3х кратное введение, частое проведение повторный курсов иммунизации. Убитые корпускулярные вакцины обладают выраженной токсичностью и аллергенностью. Их важнейшее преимущество по сравнению с живыми -они никогда не вызывают инфекционных заболеваний.
Вакцины для плановой вакцинации детей: АКДС, БЦЖ, АКаДС, ИПВ (инактивированная полиомиелитная вакцина).
После вакцинации развивается искусственный активный иммунитет. Возникает после контакта с антигеном. Это предусматривает реализацию всех этапов иммунной реакции с формированием антиген-специфических Т-лимфоцитов и антител, а также сохранением иммунной памяти. Преимуществами активного иммунитета является более высокая эффективность, так как наработанные иммунные факторы наиболее соответствуют конкретному антигенному раздражителю, а также длительный, а порой и пожизненный, эффект последействия, связанный с формированием клеток памяти и поддержанием защитного титра специфических антител в сыворотке крови и других биологических жидкостях организма. К недостаткам активного иммунитета следует отнести значительную энергозатратность и сравнительно длительный срок развития. Не передается по наследству.
| 27. Вакцины. Основные требования, предъявляемые к вакцинам. Живые вакцины: аттенуированные, дивергентные (БЦЖ, вакцина против натуральной оспы), рекомбинантные. Принципы получения, примеры. Характеристика поствакцинального иммунитета. ОТВЕТ: Живые вакцины готовят из вакцинных штаммов бактерий, риккетсий, вирусов, полученных различными методами селекции. Вакцинные штаммы являются аттенуированными, сохранившими незначительную остаточную вирулентность и не способны вызывать клинически выраженную инфекцию. Их получают путем снижения вирулентности при культивировании в неблагоприятных условиях (при пониженной или повышенной температуре, на питательных средах с определенными добавками) или путем пассажей на маловосприимчивых животных, в куриных эмбрионах и клеточных культурах, выделением аттенуированных мутантов от больных или из внешней среды, воздействием мутагенов. В аттенуированных штаммах инактивированы или репрессированы гены отвечающие за вирулентность. В связи с тем, что живые вакцины способны вызывать вакцинную инфекцию (живые аттенуированные микробы размножаются в организме, вызывая воспалительный процесс проходящий без клинических проявлений), они всегда вызывают перестройку иммунобиологического статуса организма и образование специфических антител. Это так же может являться недостатком, т. к. живые вакцины чаще вызывают аллергические реакции. Вакцины данного типа, как правило, вводятся однократно. Примеры: сибиреязвенная вакцина, чумная вакцина, бруцеллёзная вакцина, БЦЖ вакцина, оспенная дермальная вакцина. - аттенуированные - препараты, действующим началом которых являются ослабленные тем или иным способом, потерявшие вирулентность, но сохра- нившие специфическую антигенность штаммы патогенных микроорганиз- мов (бактерий, вирусов), получившие название аттенуированных штаммов. - дивергентные - получают на основе непатогенных штаммов микроорга- низмов, имеющие общие протективные антигены с патогенными для чело- века возбудителями инфекционных болезней (вакцина против натуральной оспы человека - используется вирус оспы коровы, вакцина БЦЖ - использу- ются микобактерии бычьего типа). - рекомбинантные - на основе получения непатогенных для человека ре- комбинантных штаммов, несущих гены протективных антигенов патоген- ных микробов и способных при введении в организм человека размножать- ся, синтезировать специфический антиген и создавать иммунитет к патоген- ному возбудителю. Поствакционный иммунитет (искусственный активный) клеточный или гуморальный по своей напряженности приближается к постинфекционному. |
28. Вакцинация. Эффективность вакцинации. Национальный календарь прививок РФ: цель проведения вакцинации детей и подростков, характеристика вакцин.
ОТВЕТ:
Эффективность вакцинации зависит от биологических свойств возбудителей и изготовленных из них препаратов, способов введения вакцин и иммунореактивности макроорганизма.
Вакцинных препараты могут вводиться в организм человека парэнтерально (внутримышечно, подкожно, в скарифицированную кожу), перорально, интраназально, а также в свечах и клизмах.
Для выработки прочного и длительного иммунитета необходим достаточным контакт макроорганизма и антигена, поэтому во многих случаях применяется повторенная вакцинация, сроки очередного введения вакцины зависят от свойств данного биопрепарата. Требуется определенный период времени для развития гуморального или клеточного иммунитета. Не у всех вакцинированных лиц возникает достаточная степень невосприимчивости, у некоторых людей иммунореактивности снижена, может развиваться иммунодефицитное состояние. Эффективность вакцинации зависит от типа и качества применяемой вакцины и способности возбудителя вызывать стойкий иммунный ответ. Согласно национальному календарю прививок РФ всем детям в обязательном порядке проводится вакцинация против 10 но нозологических форм -туберкулез, вирусный гепатит В, дифтерия, коклюш, столбняк, полиомиелит, корь, краснуха, эпидемический паратит, грипп. Против 13 нозологических форм только по эпидемическим показаниям- туляремия, чума, бруцеллез, сибирская язва, бешенство, лептоспироз, клещевой энцефалит, лихорадка Ку, желтая лихорадка, брюшной тиф, менингококковая инфекция, вирусный гепатит А, холера.
ОТВЕТ:
Анатоксины нередко относят к молекулярным вакцинам. Получают из бактериальных экзотоксинов путем 3-5 недельного воздействия 0,3-0,4% формалина при температуре 37-40 С. При совместном воздействии этих факторов экзотоксин теряет свою ядовитость, сохраняет антигене и иммуногенные свойства. Полученные анатоксины подвергают очистке от балластных веществ(питательной среды, продуктов метаболизма), концентрируют и адсорбируют на гидроксиде алюминия, что повышает иммуногенность. У анатоксинов относительно низкая реактогенность, поэтому мало противнопоказаний к применению. Очищенные адсорбированные анатоксины выпускают в жидком виде, применяют для создания анти токсического иммунитета при таких инфекциях как дифтерия, столбняк, газовая анаэробная инфекция, ботулизм, стафилококковая и синегнойная инфекция и др. , возбудители которых выделяют экзотоксины, играющие первостепенную роль в патогенезе заболевания.
АНТИГЕННАЯ ЕДИНИЦА (АЕ), единица измерения антигенных и антитоксич. свойств токсинов и анатоксинов. По Рамону, характеризуется величиной I/Lf, к-рая соответствует кол-ву антитоксич. единиц, полностью связывающих 1 мл токсина или анатоксина. По Гленни, АЕ измеряется Lf-limes floculation (порогом флоккуляции), т. е. миним. дозой анатоксина, полностью связывающей одну антитоксич. единицу антитоксина.
АССОЦИИРОВАННЫЕ ВАКЦИНЫ (живые + инактивированные) комплексные вакцины представляют собой комплекс различных типов вакцин и предназначены для одновременной иммунизации против разных инфекций. Отдельные компоненты такой вакцины должны быть взяты в дозировках не создающих конкуренции, чтобы иммунитет формировался ко всем антигенам с одинаковой интенсивностью.
-поливакцина - содержит однородные антигены (полиомиелитная - типы I, П, III; полианатоксины).
- комбинированные - состоят из разнородных антигенов (АКДС-вакцина).
Возникает искусственный активный иммунитет
30. Диагностические сыворотки: подразделение, состав, принципы получения. Агглютинирующие сыворотки. Отличия адсорбированных и неадсорбированных агглютинирующих сывороток. Практическое применение. Примеры.
ОТВЕТ:
Диагностические сыворотки содержат известные антитела, их применяют в серологических реакциях для определения выделенного возбудителя инфекционного заболевания или его токсина, а также доя выявления неизвестных антигенов непосредственно в исследуемом материале или из объектов окружающей среды. Их получают путем многократной иммунизации (гипериммунизации) животных (кроликов, баранов) различными антигенами -взвесью микробов или выделенными из них и очищенными микробными антигенами, анатоксинами, чужеродными сывороточными белками и другими корпускулярными и растворенными антигенами.
Диагностические сыворотки под разделяют в зависимости от реакции на агглютинирующие (адсорбированные и неадсорбированные), преципитирующие, гемолитические и антитоксические.
Агглютинирующая сыворотка - сыворотка крови животных, способная вызывать реакцию агглютинации, получаемая при иммунизации животного определённой бактериальной культурой.
Неадсорбированные и адсорбированные агглютинирующие сыворотки.
Неадсорбированные (видовые, нативные) сыворотки обладают высоким титром, но недостаточно специфичны. Видовые сыворотки содержат несколько типов антител соответственно набору антигенов того вида бактерий, которым проводилась иммунизация животного. Могут содержать групповые антитела за счет которых происходит агглютинация не только с гомологичными бактериями но и с родственными бактериями. Часто групповых агглютинация встречается у представителей рода Salmonella. Для того чтобы избежать групповой агглютинации применяют адсорбированные монореципторные сыворотки. Они характеризуется низким титром, их применяют в реакциях агглютинации на стекле. Для устранения всех групповых антител сыворотку последовательно инкубируют с разными видами родственных микроорганизмов.
Адсорбированные сыворотки применяют при идентифика ции выделенных возбудителей в реакции агглютинации на стекле (пластинчатый метод).
Агглютинирующие сыворотки наиболее широко применя ются при диагностике заболеваний, вызываемых бактериями семейства Enterobacteriaceae. Так, при идентификации эшерихий используются поливалентные и типовые ОК-сыворотки; при дифференциации сальмонелл - набор сывороток: агглю тинирующая адсорбированная поливалентная сальмонеллезная О-сыворотка (групп А, В, С, Д, Е) - для определения принадлежности к роду Salmonella, при положительном ре зультате - определяют отдельно с каждой сывороткой (входя щей в смесь) серологическую группу и в заключение опреде ляется серологический тип выделенного возбудителя с моно-рецепторными Н-сыворотками сальмонелл, входящих в данную группу.
Раздел Вирусология. Вопрос 4
Репродукция вирусов в клетке (продуктивная инфекция ) – единый процесс, который условно подразделяют ан несколько этапов. Начальный 3 этапа являются подготовительными. Собственно репродукция начинается с 4 этапа. Процессы репродукции у разных семейств и родов вирусов имеют существенные отличия. Но в общих чертах происходят следующие закономерности:
1. Адсорбция вирионов на клетке осуществляется при наличии специфических рецепторов. У простых вирусов это прикрепительные белки на поверхности капсида, а у сложных – гликопротеины, образующие шипики на поверхности суперкапсида. Со способностью вирусов прикрепляться к различным клеточным рецепторам связан тропизм вирусов.
2. Проникновение вириона в клетку
Путем рецепторного эндоцитоза – в месте адсорбции вируса образуется эндосома, содержащая вирус. Она объединяется с клеточной лизосомой и вакуолью, образуя рецептосомому. (простые и сложные вирусы)
Слияния мембран суперкапсида вируса и клетки. Нуклеокапсид оказывается в цитоплазме (сложные вирусы)
Возможно сочетание этих двух процессов.
3. Депротеинизация («раздевание») вирусов с целью высвобождения нуклеиновой кислоты, которая приобретает способность индуцировать репродукцию вирусов.
4. Экспрессия вирусного генома. Начинается с транскрипции (образование комплементарных и-РНК). Затем идет трансляция (синтез белка) на клеточных рибосомах. Моноцистронные и-РНК кодируют отдельный белок, полицицистронные и-РНК длинный общий белок, который нарезается клеточными протеазами на отдельные белки.
А) У вирусов с двунитевой ДНК: геномная ДНК – транскрипция – и-РНК- трансляция – белок. Если происходит в ядре, то работает клеточная ДНК-зависимая-РНК-полимераза, если в цитоплазме, то вирусная транскриптаза. Последовательно кодируются ранние и-РНК, затем поздние и-РНК, следовательно, сначала происходит трансляция сначала ранних неструктурных белков, затем поздних структурных. Такими вирусами является вирус гепатита В, вирус герпеса, вирус папилломы.
Б) У плюс-РНК геномная РНК является сразу информационной РНК, поэтому: Геномная плюс-РНК - трансляция – белок.
В) У минус- РНК6 Геномная минус-РНК-транскрипция-и-РНК-трансляция-белок.
Г) у ретровирусов: Геномная РНК-комплементарная ДНК-транскрипция-и-РНК-трансляция-белок.
Затем идет репликация вирусного генома (синтез множестав копий нуклеиновой кислоты)
А)ДНК-геномы клеточной ДНК-зависимой ДНК-полимеразы
Б) Плюс-РНК вирусиндуцированной РНК-полимеразы
В)Минус-РНК с помощью РНК-зависимой РНК-полимеразы.
Г) у ретровирусов клеточной ДНК-зависимой РНК-полимеразы. Для них необходима интеграция ДНК провируса в хромосому клетки, а также сочитание интегративной и продуктивной инфекции.
Дизъюктивный (разобщенный) способ репродукции- синтез нуклеиновых кислот и белков вирусов в разных частях клетки и неодновременно.
5. Формирование вирионов (в цитоплазме). Простые вирусы путем самосборки, образуя нуклеокапсид. Сложные вирусы сначала образуют нуклеокапсид, затем взаимодействуют с модифицированной клеточной мембраной, одеваясь суперкапсидом и формируя М-слой.
6. Выход вирионов из клетки при разрушении, лизисе или путем почкования(для сложных вирусов, которые при этом одеваются суперкапсидом)
Суть механизма РНК-интнрференции заключается в том, что при введении в клетки короткой двунитевой РНК, она способна вызывать специфическое разрушение той мРНК, с которой имеет гомологию. Двухцепочная РНК при этом распадается на короткое фрагменты, обозначаемые как малые интерферирующие РНК. Это используется для подавления распространения инфекций.
Инфекционность вирусов способна реализоваться в восприимчивых клетках, обладающих пермиссивностью. Только пермиссивные клетки способны обеспечить все этапы репродукции(продуктивная инфекция ). Существуют непермисссивные и полупермиссивные клеточные системы, способные воспринять вирус, но не обладающие всеми факторами, обеспечивающими их репродукцию. В этом случае возникает абортивна я инфекция без образования вирусного потомства.
Вирусная инфекция на уровне клетки бывает автономной или интегративной. Автономная- продуктивная инфекция, при которой вирусный и клеточный гномы, находясь в одной клетке взаимодействуют друг с другом, но при этом остаются обособленными. В противоположность этому, при интегративной инфекции вирусный геном объединяется с клеточным геномом, становясь его частью(для ДНК-вирусов). Это может привести к нарушению синтеза белка в клетке, к неконтролируемому делению клеток-трансформаци и и развитию опухоли.
Инаппарантная форма инфекции - острая бессимптомная инфекция, протекающая при полном отсутствии клинических и биохимических признаков. Обнаруживается лишь закономерное динамикой маркеров острой вирусной инфекции.
Для сохранения вируса как биологического вида очень важной является способность многих вирусов персистировать в клетках организма, тоесть сохраняться в организме длительное время, иногда пожизненно, периодически выделяясь в окружающую среду. Активируясь, вирусы могут вызывать эндогенную инфекцию (при недостаточности иммунитета) . в результате может развиться хроническая инфекцию.
Вирусная инфекция может быть цитолитической (клетка погибает, лизируетсяпосле 1-го цикла репродукции) и нецитолитической (клетка продолжает функционировать какое-то время и осуществлять репродукции вирусов в несколько циклов, образуя симпласты; внутриклеточные включения; возможна латентная инфекция , когда имеются только функциональные нарушения, а их видимое проявление отсутствует; возможна трансформация клеток и опухолевой рост.
Для медленных вирусных инфекций характерны особенности: длительный инкубационный период (от нескольких месяцев до десятилетий; медленное развитие заболевания(без ремиссий); преимущественное поражение ЦНС; неизбежный смертельный исход. Группы медленных инфекций:
1. Вызываемые при определенных условиях обычными, «каноническими» вирусам после перенесенных вирусных инфекций(корь, краснуха, клещевой энцефалит)
2. Прионные вирусные инфекции. Прионы - инфекционные вирусные белки, которые являются видоизмененными клеточными белками, образовавшимися вследствие мутаций в кодирующем эти белки гене.
Раздел Вирусология Вопрос 5